First Skinnerian genes discovered
New molecular learning mechanism in fruit flies
Press |
Press coverage running tab: Radio shows covering the research: This Week in Science, Deutsche Welle (English) |
This is my attempt at explaining the content of our publication entitled "Double Dissociation of PKC and AC Manipulations on Operant and Classical Learning in Drosophila" to a more general audience, so please forgive my simplifications and omissions. Please do not hesitate to contact me for any further clarification.
Summary
We have discovered the first learning genes specific for operant conditioning in the fruit fly Drosophila. Our research, published July 31, 2008 in the journal Current Biology, may help understand the molecular processes underlying addiction. We tested genetically engineered flies, tethered to fly stationarily in a computer-controlled environment in three different learning paradigms and found at least two basic molecular learning mechanisms. It is still very early, but it is possible that these two processes are sufficiently separate that they could take place concurrently in the same nerve cells without interference.
Background
While the Russian physiologist I.P. Pavlov trained his dogs to salivate to the tone of a bell by feeding them immediately after he rang the bell (Pavlovian or classical learning), American psychologist B.F. Skinner trained pigeons and rats to perform a behavior for a food reward (operant or instrumental learning). For his experiments, Skinner devised experimental chambers, later called Skinner-boxes, in which e.g. a rat had to press a lever in order to obtain food pellets (see video).
Ever since these forms of learning were established, people have debated as to how the two relate to each other. After all, the lever might signal food to the rat just as the bell signals food for the dog and both learn the same thing, despite all the differences in the procedures.
To study the mechanisms underlying learning, modern neurobiological resarch today concentrates mainly on the up- or downregulation of the connections between nerve cells ('synaptic plasticity'). This concentration makes a lot of sense because synaptic plasticity appears to be involved in most if not all learning tasks and it is sufficient to change the strength of synaptic connections to store information in a neural network. Many of the genes regulating synaptic plasticity have originally been discovered individually in Pavlovian conditioning experiments. Only later was it discovered that they all share their participation in this wide-spread phenomenon and therefore are critical also for many other learning tasks, some of them operant. However, pilot experiments by then undergraduate student Wolfgang Plendl seemed to suggest that at least the two major forms of associative learning could be differentiated at the genetic level. He found that one of the earliest known Pavlovian learning mutants in Drosophila, rutabaga, performed well in a certain group of operant experiments and that inhibiting the enzyme protein kinase C (PKC), impaired flies in exactly the same group of experiments. Until now, PKC was not known to be involved in changing synaptic strength or learning. Inspired by Wolfgang’s pilot experiments, I set up the three experiments which I think may have solved the 80 year-old debate.
Methods
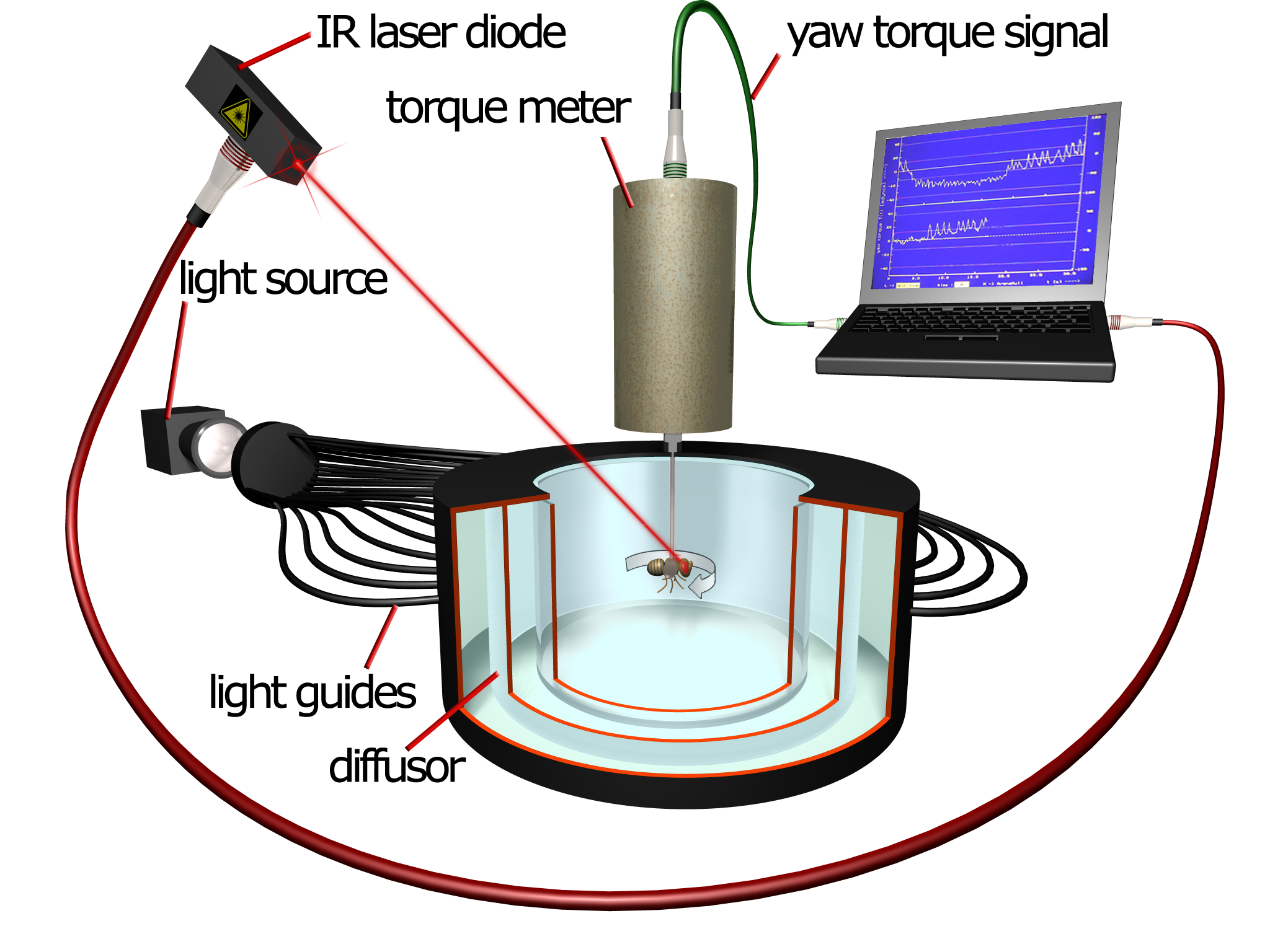
This is the setup I used:
The problem with the Skinner-boxes was that you needed the lever. Take the lever away and you have nothing for the animal to learn. So we had to use an experiment where the animal could do something without changing anything in its environment. This seemingly impossible experiment can be done in the setup shown in the video above. The flies are attached to a measuring device which measures the fly’s attempted turning maneuvers. Under the control of a computer, a punishing heat beam and/or initially neutral colors can be made to follow, say, either right or left turning attempts.
We used this machine to set up three experiments. Two which were designed to be similar to the standard Pavlovian and Skinnerian experiments, and a third one which was designed to mimick taking the lever out of the box to study the behavior in isolation.
In the first of the three experiments, the turning attempts of the fly were irrelevant and only the color signaled the heat, similar to the bell for Pavlov’s dogs. In the second experiment, turning attempts in one direction switched both heat off and changed the color stimulus, similar to Skinner’s original experiment with the rats pressing the lever for food. In the final experiment, no color was presented and only turning attempts in one direction switched the heat off. Clearly, the third experiment was the critical one, because removing the colors was supposed to be analogous to removing the lever in a Skinner-box. Which of the genetically engineered flies would fail in this paradigm?
 |
Experiment 1 - Only classical cues: The fly controls the angular position of a drum with four identical vertical bars in a flight simulator-like situation. The coloration of the arena is switched between bars, such that flying towards one pair of opposing bars leads to green coloration and towards the other pair to blue coloration. During training, heat is made contingent on one color, irrespective of the turning maneuver which changed flight direction. The colors are analogous to the bell in Pavlov's original experiment. |
 |
| Experiment 2 - operant and classical cues combined: Throughout the experiment, one yaw torque domain is coupled to one color and the other to the other color (e.g., right turning causes green illumination and left turning blue illumination of the environment). During training, heat is made contingent on one of the two yaw torque/color combinations. This experiment was designed to be analogous to Skinner's original experiment where rats have to press a lever for a reward (see video above): The yaw torque/color combination is analogous to the pressing/lever combination. |
 |
| Experiment 3 - only operant cues: There are no visual cues for the fly. During training, heat is made contingent on either left- or right-turning yaw torque. The turning attempts of the fly correspond to the lever-pressing of Skinners rats, with the important difference that in this experiment, there is no lever which could serve as an environmental (i.e., classical) signal for the reinforcement. |
Results
The results were surprisingly clear cut: the two experiments most similar to the original experiments of Pavlov and Skinner (i.e., experiment one and three) both required the well-known learning gene rutabaga. Thus, these experiments confirmed the last decades of research which have shown similar mechanisms for both kinds of experiments. The one experiment which was impossible to do in rats or other mammals turned out to be genetically very different: rutabaga-mutant flies performed even better than normal flies, and PKC-inhibited flies did not learn even though they performed well in the first and third experiment. Apparently, when the flies have the opportunity to learn both about their behavior and the external stimuli, learning about the external stimuli dominates the whole process. This indicates that the reason why most people thought there was only a single genetic network underlying so many forms of learning, may be that these forms of learning all involve learning about your environment.
Here is a table to graphically illustrate the results, red means involved, green not involved:
rutabaga |
PKC |
|
| Pavlovian (exp. 1) | ||
| Skinnerian (exp. 2) | ||
| Pure operant (exp. 3) |
Discussion
Apparently, as soon as predictive stimuli are present in nominally operant learning situations, not only do these stimuli become equivalent to Pavlovian stimuli with respect to their independence from the behavior with which they were learned (shown in an earlier study), but these �composite� experiments also cannot be distinguished genetically from Pavlovian experiments any more. The procedural classification of learning experiments into operant and Pavlovian is not reflected in the biological processes taking place in the animal. Maybe the old textbook dichotomy of associative learning being neatly divided into operant and Pavlovian conditioning will have to be revised.
There are now several lines of evidence which suggest that this new, PKC-dependent form of behavioral learning is akin to habit learning, which is used to study addiction in mammalian model systems. Therefore, I am also pretty excited that my colleagues in my old lab in Houston, Texas have just now found out that PKC is also involved in operant learning in the marine snail Aplysia. Their research will appear in the September 11 issue of the journal Neuron and will present quite a lot more detail on the other components of the molecular network underlying operant learning. Our combnied findings raise the possibility that just like synaptic plasticity, the new PKC-dependent learning mechanism may also be evolutionary conserved from snails and flies to mammals. At this point, nobody knows what kind of mechanism this may turn out to be: another type of synaptic plasticity or some form of neural plasticity. In any case, the independent simultaneous discovery of of a novel, PKC-dependent learning mechanism in two different invertebrate model systems makes a compelling case that this research might turn out to be very relevant for human brain resarch as well.
The next steps will be to examine if the overtraining procedures known to lead to habit learning in mammals also lead to behavioral learning in fruit flies even when the color is signaling heat punishment, as well as which structures in the fly brain require which isoform of PKC in order for the flies to learn about the consequences of their behavior. In Drosophila, there are six genes coding for a total of 10 different isoforms of PKC (all of which appear to be inhibited by our method), so this task will take some time.
The research will appear online on July 31, 2008 on the website of the journal Current Biology. Press embargo in effect until Thursday, July 31, 2008, 12:00 Noon - Eastern Time (U.S.).
The general experimental procedures we use for all our Flight Simulator learning experiments are described in this JoVE video:
Brembs and Plendl (2008): Double Dissociation of PKC and AC Manipulations on Operant and Classical Learning in Drosophila, Current Biology, 18(15) 1168-1171, doi:10.1016/j.cub.2008.07.041
Manuscript summary:
Learning about relationships between stimuli (i.e., classical conditioning) and learning about consequences of one's
own behavior (i.e., operant conditioning) constitute the major part of our predictive understanding of the world. Since these forms of learning were recognized as two separate types 80 years ago, a recurrent concern has been the
issue of whether one biological process can account for both of them. Today, we know the anatomical structures required for successful learning in several different paradigms, e.g., operant and classical processes can be localized to different brain regions in rodents and an identified neuron in Aplysia shows opposite biophysical changes after operant and classical training, respectively. We also know to some detail the molecular mechanisms underlying some forms of learning and memory consolidation. However, it is not known whether operant and classical learning can be distinguished at the molecular level. Therefore, we investigated whether genetic manipulations could differentiate between operant and classical learning in Drosophila. We found a double dissociation of protein kinase C and adenylyl cyclase on operant and classical learning. Moreover, the two learning systems interacted hierarchically such that classical predictors were learned preferentially over operant predictors.




