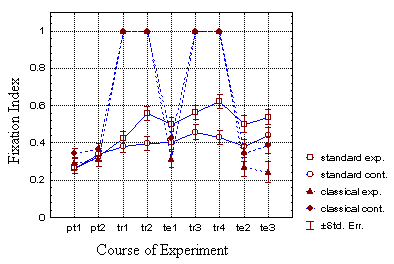
Fig. 27: Mean fixation indices of all four groups. Note that the training indices of the classical groups are always 1.0 due to the static presentation of the patterns.
In both operant groups fixation increased during the course of the experiment, whereas in the classical groups only the control group showed an increase in fixation (Fig. 27).
Fig. 27: Mean fixation indices of all four groups. Note that
the training indices of the classical groups are always 1.0 due to the
static presentation of the patterns.
Compared to the operant group, the flies that were presented with static patterns fixated the patterns significantly worse at t2 (periods 8 and 9; Table 4), whereas there was no difference at t1 (Fig. 27). Since the two respective control groups do not reveal any significant differences in fixation, this effect has to be attributed to the different reinforcement procedures. For the same reasons, the shift in the amount of total arena rotation at t2 has to be due to the training procedures (Table 4). Classical and operant groups showed no difference in the number of quadrant changes (Fig. 28).
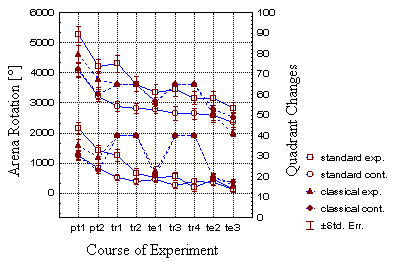
Fig. 28: Combined presentation of total amount of arena rotation
(upper four lines, left abscissa) and number of quadrant changes per period
(lower four lines, right abscissa). Note the fixed values for the classical
training periods.
Decreasing numbers of quadrant changes and amount of arena rotation (Fig. 28), together with decreasing spike numbers support the view that the flies become acquainted with their situation and calm down during the course of the experiment. The force of the body saccades, however, remains constant throughout the experiment (Fig. 13).
The finding that the operantly trained flies fixate best confirms earlier experimental data from Dill et al. (1995) where it was demonstrated that flies prefer flight directions in the middle of the 'cold' quadrants when they have been heated in the other sectors. It seems as if this increase in fixation is an acquired behavioral strategy and not the consequence of decreasing overall activity (see below, 3.6). While at t1 the fixation index is weakly (about -0.2) but significantly correlated with spike number and total amount of arena rotation (i.e. activity) this correlation ceases at t2. It is therefore not surprising that the flies that show the highest fixation index at t2 generate more spikes and cause more arena rotation than the others (however, this group was already more active at t1).
If fixation were an actively selected behavioral strategy, the poor fixation in the classically conditioned flies might be due to the avoidance of the 'hot' pattern orientation (i.e. anti-fixation) while fixation in the 'cold' sectors is within control limits. This view is discussed more thoroughly below (3.6).
| standard | s-control | classic | c-control | |
| FixInd*** | 0.52 | 0.41 | 0.26 | 0.37 |
| Spikes* | 35.4 | 30.5 | 27.7 | 33.9 |
| SpDur** | 0.48s | 0.48s | 0.54s | 0.49s |
| ARot* | 2986° | 2470° | 2285° | 2658° |
Table 4: Values for those variables, where the null hypothesis that the four groups were drawn from one population could be rejected, averaged over the two last periods (t2). FixInd - fixation index; Spikes - number of spikes per two minute period; SpDur mean spike duration, ARot - total amount of arena rotation. * - p<0.05; ** - p<0.01; *** - p<0.001.
It cannot be ruled out, however, that the poor fixation is due to fatigue
caused by the high amount of heat each fly is receiving during classical
training: the flies get weaker as they are dehydrated in the course of
the training. Consequently, they generate less and smaller spikes (Figs.
12 and 13) as well as a reduced amount of optomotor balance (quiet mode,
Fig. 8). Heisenberg and Wolf (1979) report that 'non-fixation' is
particularly common in flies using the quiet mode of flight:
"[...] the stripe may suddenly start to be shifted to
any position and may be kept there quietly for some time. We call this
behavior 'non-fixation'."
Picturing this quiet mode might also explain the extremely long
spike duration in the classical test group: it leads to a later reversion
of the torque slope after each spike which is the confinement of spike
duration as defined above (Fig. 10). Consequently, spike duration is negatively
correlated with arena rotation, spike number and spike amplitude (not shown).