As discussed above (3.5.2), the flies primarily modulate two polarity
independent characteristics of turning maneuvers in order to avoid certain
flight directions in the flight simulator: spike amplitude and spike timing.
Two sources of evidence support this view: the comparison of the test values
with the respective spontaneous values generated by the control flies (see
above, 3.5.2) and a correlational analysis in all four groups with the
preference values (Table 5).
standard
s-control
classic
c-control
AmpInd8
0.60***
0.60***
0.40***
0.45***
AmpInd9
0.60***
0.68***
0.46***
0.46***
LatInd8
0.09
0.45***
0.15
0.14
LatInd9
0.40***
0.25*
-0.01
0.14
ISIInd8
0.79***
0.68***
0.72***
0.83***
ISIInd9
0.76***
0.90***
0.83***
0.79***
NumInd8
0.56***
0.70***
0.62***
0.76***
NumInd9
0.75***
0.75***
0.61***
0.84***
Table 5: Spearman Rank Order Correlations with the preference
index at t2. AmpInd - amplitude index, LatInd - latency index,
ISIInd - ISI index, NumInd - number index. * - p<0.05; ** - p<0.01;
*** - p<0.001.
The only obvious difference between the operant and the classical groups
to be detected in Table 5 is in the latency indices. This is not surprising,
since only one spike is counted for each stay in one quadrant while the
fly is generating many more afterwards and therefore has many more options
to choose flight direction. Small disturbances in flight behavior due to
rather long phases of open loop might exert a large effect in such a variable.
Furthermore, even in those cases where the correlation is significant,
the values are lower than all the other correlations. From the data presented
so far one can infer that flies that have high preference indices produce
more and larger spikes in the 'hot' than in the 'cold' sectors, even if
the heat is switched off, regardless of their training procedure. Performing
a correlational analysis among the different parameters yields results
that strengthen this view: those flies that generate more spikes in the
'hot' sectors do also tend make them larger (significant positive Spearman
Rank Order Correlations at t2 between the ISI index, the number
index and the amplitude index in all four groups, data not shown). This
is also the case for spontaneous behavior (see table 5): the flies use
the same motor-output to express their endogenous preference for a certain
flight direction. This in turn leads to an important corollary: both training
procedures might modulate the endogenous preference of each fly.
Another piece of evidence pointing in this direction are some of the
variables measured independently of quadrant quality (spike numbers, fixation
index and quadrant changes): in the test groups they are significantly
correlated with the respective preference indices, whereas in the control
groups they are not (Table 6). Does this mean that flies that are calmer
and fixate better are also better learners?
standard
s-control
classic
c-control
Spikes8
-0.27**
-0.01
-0.27**
-0.10
Spikes9
-0.04
0.02
-0.14
-0.20
FixInd8
0.53***
-0.10
0.34***
0.17
FixInd9
0.36***
-0.15
0.34***
0.11
QuCh8
-0.48***
0.06
-0.47***
-0.19
QuCh9
-0.23*
0.00
-0.33***
-0.16
Table 6: Spearman Rank Order Correlations with the preference
index at t2. Spikes - number of spikes per two minute period;
FixInd - fixation index; QuCh - number of quadrant changes per two minute
period. * - p<0.05; ** - p<0.01; *** - p<0.001.
This inference is not necessarily true, of course, since performing
the same analysis with the absolute preference indices yields identical
results for all four groups (not shown). In other words: all the reinforcer
does is switching the endogenous preference in all flies to the same direction.
This is mimicked by using absolute preference indices. Then of course it
becomes clear why reduced activity and better fixation leads to higher
scores: the lower the activity is, the longer the dwelling times in each
sector and the higher the preference indices - independently of whether
the fly has learned anything. This outcome would be expected if fixation
were correlated with the expression rather than with the acquisition of
memory.
Among all the variables measured independently of the differently treated
(hot/cold) sectors, the null hypothesis that all groups were still from
the same population had to be rejected in four variables: fixation index,
spike number, spike duration and total amount of arena rotation (Table
4). Interestingly, the fixation indices indicate that the flies trained
in closed loop fixate the pattern more closely to the very front (i.e.
generate more spikes towards the pattern) than do the flies presented stationary
patterns during training. As discussed above (3.5.4), this effect is largely
due to strong deviations among the two test groups (p<0.001), whereas
the control groups do not differ (p=0.449).
After evaluating a number of spike polarity dependent variables, only
the number of spikes towards the pattern compared to the number of spikes
away from the pattern yielded results that could be related to learning
(Table 7).
standard
s-control
classic
c-control
PolHot8
-0.49***
-0.30**
-0.34***
-0.27**
PolHot9
-0.41***
-0.50***
-0.48***
-0.43***
PolCold8
0.49***
0.42***
0.31**
0.49***
PolCold9
0.51***
0.40***
0.20
0.28**
PolLat8
0.02
0.17
0.11
0.11
PolLat9
0.05
0.19
0.02
0.14
Table 7: Spearman Rank Order Correlations with the preference
index at t2. PolHot - 'hot' polarity index; PolCold - 'cold'
polarity index; PolLat - polarity latency index. * - p<0.05; ** - p<0.01;
*** - p<0.001.
As fixation index and overall polarity index are of course highly correlated
(mean Spearman Rank Order Correlation at t2 0.46, p<0.001),
the polarity indices in the differently treated sectors are especially
telling: In the 'cold' sectors, more spikes towards the pattern were generated
than away from the pattern and vice versa in the 'hot' quadrants.
However, there was no 'training effect' i.e. unproportionally large values
for the training periods, indicating that this behavior is largely independent
from the reinforcer. This was the case for all the variables connected
with spike polarity (not shown).
In the light of spike polarity, the poor fixation of the classical test
group compared to the operant test group might indeed reflect different
behavioral strategies acquired by the different training procedures: during
operant training the flies learn that the centers of the 'cold' quadrants
are 'safe' (Dill et al. 1995). During classical training, the sector borders
are not perceptible for the fly - the flies are punished with the pattern
in the centralmost position. They might even learn to avoid this central
position. This experience might be more salient to the fly than the unpunished
position of the other pattern orientation. The data on spike polarity points
in this direction: while in the 'cold' sectors the polarity index of the
classical test group does not rise above the control level as does the
operant group, the drop in the 'hot' sectors is significant and indistinguishable
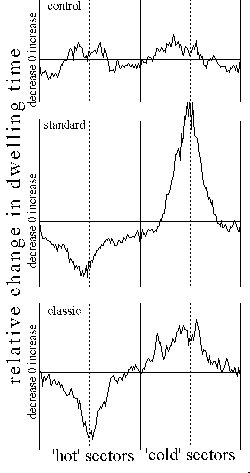
from the operant group. Fig. 29 illustrates this accurately: in the 'hot'
quadrants both groups equally decrease fixation (i.e. decrease the number
of spikes towards the pattern), while in the 'cold' sectors only the operant
group decreases its stays near the borders and increases fixation. The
flies in the classical group increased dwelling times in the entire 'cold'
sector.
Fig. 29: Illustration of the changes in dwelling time distribution.
From the position trace, a dwelling time/position histogram for 'hot' and
'cold' quadrants is calculated for both pre-test (t1, periods
1 and 2) and test (t2, periods 8 and 9). The difference between
these histograms at t1 and t2 is depicted here. Dotted
lines indicate the centers of the patterns, vertical lines quadrant borders.
The horizontal lines depict zero change in dwelling time from t1
to t2.
This difference is also reflected in the results of a correlational
analysis among the measured behavioral parameters: while spike polarity
at t2 in the 'hot' quadrants was significantly correlated (negatively)
with indices describing modulation of polarity independent spike parameters
in all four groups, spike polarity in the 'cold' sectors was not correlated
with the other indices in the classical group (Table 8).
standard
s-control
classic
c-control
AmpIndH
-0.18
-0.29**
-0.27**
-0.11
ISIIndH
-0.43***
-0.45***
-0.36**
-0.22*
NumIndH
-0.23*
-0.32**
-0.30**
-0.30**
AmpIndC
0.23*
0.34***
0.14
0.10
ISIIndC
0.44***
0.32**
0.21
0.43***
NumIndC
0.33**
0.36***
0.14
0.31**
Table 8: Spearman Rank Order Correlations with spike polarity
at period 8. Superscripts indicate with wich polarity index the variable
was correlated: H - 'hot' polarity index. C - 'cold' polarity index. AmpInd
- amplitude index, ISIInd - ISI index, NumInd - number index. * - p<0.05;
** - p<0.01; *** - p<0.001.
The modulation in the polarity of the first spike was negligible: a
mean decrease of 0.06 in the probability that the first spike is towards
the pattern in the 'hot' compared to the 'cold' sectors was the largest
value obtained (standard experiment). The overall probability was over
0.8 that a first spike in any quadrant is "towards pattern".
Summarizing the data concerning the direction of turning maneuvers,
it seems as if modulation of spike polarity is one but not a primary behavioral
strategy for orientation in the Drosophila flight simulator. 'Simpler'
strategies such as modifying the force or the frequency of the body-saccades
seems to account for more of the avoidance behavior than directed orientation
in space.