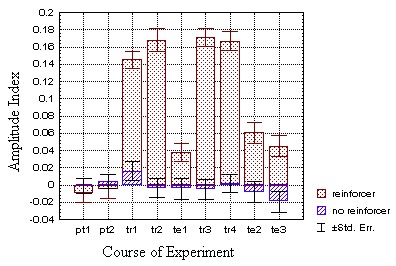
While in neither of the groups spike duration was modulated in response
to heat (data not shown), operantly trained Drosophila exhibited
a small but highly significant modulation of spike amplitude, even when
the heat was switched off during the last two periods (p<0.001, fig.
20A).
A
B Fig. 20: Mean amplitude indices for all four experimental
groups (N=100 flies each). The flies learn to generate larger torque spikes
in quadrants with one of the pattern orientations, if they were reinforced
in these quadrants during the training periods (dotted bars). This
is the case for the operantly (A) and the classically (B)
trained flies. The control groups, which did not receive any reinforcement
modulated spike amplitude randomly (hatched bars).
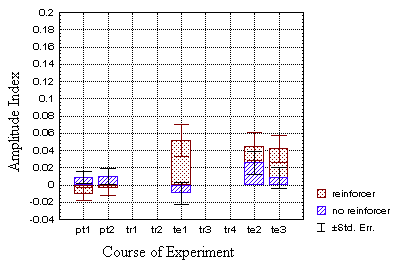
Although the classically trained flies do modulate their spike amplitude
in response to the heat as does the operantly trained group (Fig. 20B),
the difference fails to reach a level of confidence of p<0.05 at t2.
However, if one compares all three test-periods (nos. 5, 8 and 9) with
the respective control values the modulation comes to lie at a reliable
p<0.03.
The t2 amplitude indices of the classical test group show
no significant deviations from those of the operant test group (p=0.35).
Furthermore, these amplitude indices are in both groups positively correlated
with the respective preference indices at t2 (Table 5) and before
as well (data not shown). Most importantly, Drosophila seems also
to modulate the spike amplitude spontaneously: even in the control groups
is the amplitude index highly correlated with the respective preference
index (Table 5).
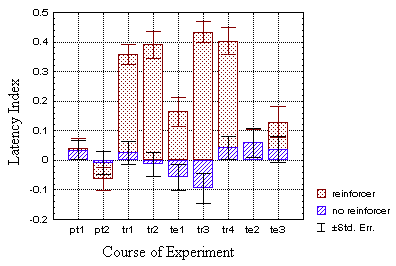
In addition to spike amplitude, the flies use the timing of spikes to
avoid the heat in the standard experiment: as noted above, often a volley
of spikes is generated to get the fly out of the heat (Fig. 21). Comparing
the spike latency in the heated sectors with that in the non-heated sectors,
shows the reaction to the heat: the time until the first spike is significantly
reduced in the 'hot' sectors (Fig. 21A).
This behavior is maintained even when the heat is permanently switched
off. This is not significant for the last two periods alone, but for all
three test periods a p<0.041 renders the effect reliable.
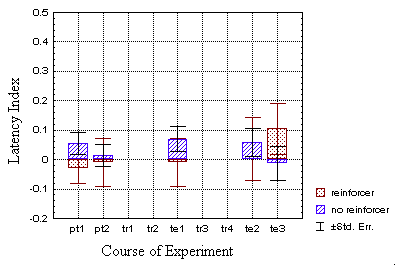
Since the latency indices of the operantly trained flies were already
very low at t2, it is not surprising that the latency indices
of all three test periods from the classically trained batch rise only
slightly above control-level (Fig. 21B). However, they could neither be
distinguished from the operant batch with a significant reliability.
A
B Fig. 21: The two test groups (dotted bars)
show mean latency indices that suggest a modulation of the reaction time
until the first spike in response to heat (see text). Most suggestive are
the high training bars in the operant group (A). In the classical
group (B), only in the last period, the first spikes are produced
quick enough in quadrants with the heated pattern orientations to bring
the index discernibly above the control group (hatched bars).
Interestingly, both groups show different correlations with the preference
indices in the respective periods (Table 5). While there is some correlation
in the operant groups - indicating that the latency until the first spike
has a certain predictive value for avoidance and learning in these flies
- there is no correlation between the latency indices and the preference
of the classically trained flies at t2. This seems to be an
effect of the open loop pattern presentation, since this is also revealed
in the classical control group (Table 5).
Even if the timing of the first spike were not modulated in response
to the heat, the fly still has the possibility to make all subsequent spikes
quicker to avoid the unpleasant flight direction. In the standard experiment,
the interspike intervals seem indeed to be shorter in heated sectors than
in 'cold' ones: flies that were able to control the appearance of the reinforcer
produce shorter interspike intervals in 'hot' sectors compared to the control
flies, even if the heat is switched off for the last two periods (t2,
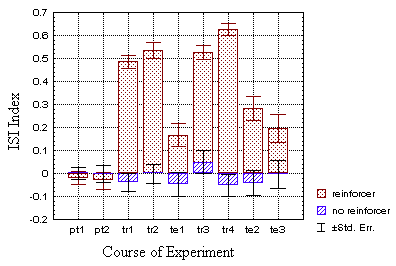
p<0.001; p=0.336 at t1; Fig. 22A).
A
B Fig. 22: Mean ISI indices for all four groups
(N=100 flies each). Drosophila produces torque spikes more quickly
when it is heated in closed loop (A, dotted bars). If the
heat is switched off, the flies still generate spikes with shorter interspike
intervals in quadrants with the 'hot' pattern orientation. The control
group, which did not receive any reinforcement only showed random modulation
of the interspike intervals (hatched bars).
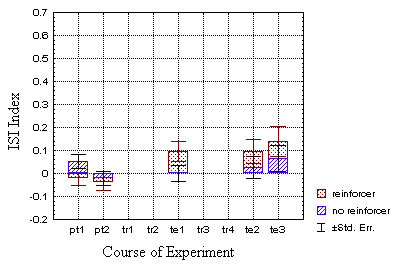
In the classical groups (B), however, statistical analysis could
not establish a significant difference between the test (dotted bars)
and the control group (hatched bars). Nevertheless, the values are
qualitatively in the proper range to assume that they perform the same
behavior as the operant group, although to a lesser degree.
Even if the plotted ISI indices of the classical test group (Fig. 22B)
seem rather well in accordance with the findings in the operant test-group,
they fail to rise above the required significance niveau. Nevertheless,
both pre-test values being in the negative range and all test-values in
the positive and above the control-values suggest a qualitatively similar
although quantitatively less strong effect. Moreover, the corrected ISI
indices at t2 (mean ISI index of the pre-tests subtracted) cannot
be distinguished statistically from the respective values in the operant
group (p>0.06).
As the amplitude index, the distance and latency indices of the two
test groups are highly correlated with the preference index at t2
(Table 5). This is also the case for the spontaneous behavior measured
in the control groups (Table 5).
Having demonstrated that at least the operantly trained flies indeed
generate spikes more quickly in those quadrants associated with the heat,
one can conclude that the spike frequency (number of spikes per time) is
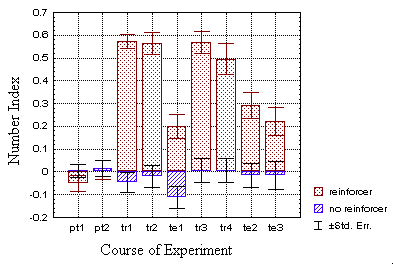
elevated as well, which can be seen in fig. 23.
A
B Fig. 23: The mean number indices in the operant groups
(A) clearly reflect the behavioral strategy of making many spikes
in previously punished flight directions: whereas the control group (hatched
bars) fails to produce other than random spike frequency modulations,
the test group (dotted bars) shows directed spike frequency modulation
in the predicted way.
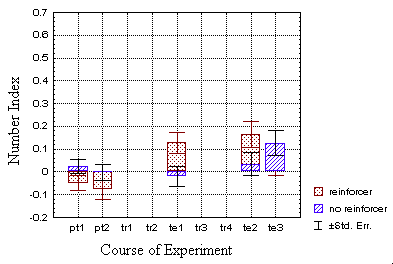
This is also visible in the classical groups (B), however to a lesser
degree (see text).
At first sight (Fig. 23B) the response of the classically conditioned
group seems to be the same as in the operant group. However, it fails to
surpass control-levels to a sufficient degree (p=0.17 for all three test
periods). However, omitting period 9 with the strikingly high control value,
the modulation exceeds the control values of periods 5 and 8 (p<0.02).
Paralleling the conditions for the interspike intervals, the number
indices of all four groups correlate with the respective preference indices
(see Table 5) and the corrected number indices in the classical test group
do not differ significantly from the values in the operant batch (p>0.05).
Taking together all the data presented so far, it seems as if the major
reason why the effects in the classical group appear to be less significant
than in the operant group, is the unusually low preference in the two pre-test
periods (t1) of the classical group, together with the unusually
high spontaneous preference in the periods 8 and 9 (t2) of the
corresponding control group. The finding, however, that the values of the
classical group are not significantly different from the operant test group
either is taken as evidence for the hypothesis that the effects are indeed
of the same nature as in the operant group. Moreover, if one expects the
control groups to exhibit symmetrical behavior, i.e. zero values in all
indices, then one can test the index values of period 8 with a Wilcoxon
Matched Pairs Test against zero. This test yields significant effects for
the number index (p<0.01, period 8) and the amplitude index (p<0.03,
period 8). The ISI index is very close to significance (p=0.067, period
8).