What do Flies Tell us about the Origins of Language?
International collaboration discovers 'language gene' necessary for fly learning
Press release (PDF): UR, FU, UM in German: UR, FU
Summary of findings
The Forkhead Box P2 (FOXP2) gene is well-known for its involvement in language disorders. We
have discovered that a relative of this gene in fruit flies, dFoxP, is necessary for a type of
learning called operant self-learning, which resembles some aspects of language
learning. This discovery traces one of the evolutionary roots of language back
more than half a billion years before the first word was ever spoken.
Intriguingly, dFoxP-function also
differentiates between self and non-self, a key process malfunctioning in autism
and schizophrenia disorders, in which FOXP2 has also recently been implicated.
Finally, dFoxP is also important for
habit formation, a common animal model for addiction.
FOXP2 and language
Even though language is so much a
part of what it means to be human, the evolution of this strikingly singular
trait is still clouded in mystery. Genetic disorders with language impairments
are a particularly effective route to uncovering the biological roots of
language. Most prominently, one mutation in the FOXP2 gene appears to affect
language acquisition in afflicted patients, without other obvious impairments (1). This gene is one of four
members of the FoxP gene family which have evolved in vertebrate animals from a
single ancestral FoxP gene by serial duplications. In invertebrates, these
duplications never took place and thus the single currently existing
invertebrate FoxP gene can serve as a model for studying the function of the
extinct, ancestral gene (Fig. 1).
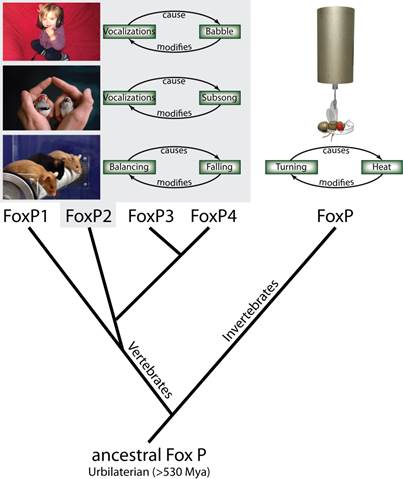
Fig. 1: Using operant conditioning to test invertebrate FoxP function.
From the single ancestral FoxP gene, four different genes have evolved in the
vertebrate lineage through serial duplications, while invertebrates have
retained a single copy of the gene. In an operant feedback loop, spontaneous
actions are followed by a given outcome as a consequence. Depending on that
outcome being desirable or not, the frequency of the action increases or
decreases in the future. For instance, vocalizations of a human infant are
followed by the perception of the resulting babbling. The deviation from the
intended articulation modifies future vocalizations until language is formed.
Similarly, in songbirds, the perceived difference between the juvenile birds (right) own
subsong and the memorized song from an adult tutor (left) modifies future
vocalizations until the species-specific adult song is produced. In mice,
balancing in the rotorod experiment is followed by eventual falling, which
provides the feedback to improve subsequent balancing movements. All three
examples have been shown to be dependent on normal FoxP2 function. Analogously,
we have tested fly FoxP function by tethering flies and measuring their turning
attempts in stationary flight. Some turning attempts (e.g. to the right) are
followed by a punishing heat beam, others (e.g., to the left) are rewarded with
turning the beam off. Continuous feedback modifies the flys turning attempts towards
the direction where the heat is off.
Studies on FOXP2 patients revealed
apraxia, i.e., the inability to articulate words and sentences, as one major
symptom. Evidence from songbirds and transgenic mouse models seems to confirm
the suspicion that the function of FoxP2 might be found in the speech component
of language (1). More than fifty years ago,
the behaviorist B.F. Skinner proposed that language might be acquired through
an operant learning process (2): the first more or less
random utterances (babbling) of infants are rewarded by their parents and
correct utterances more so than incorrect ones. Moreover, just as imitating any
movements, the ability to correctly imitate the words of others might be
inherently rewarding. Eventually, the infant learns to correctly speak the
words required to communicate their needs and affections.
Testing FoxP function in flies
Inspired by the possibility to test
for one of the evolutionary roots of language in an invertebrate animal, we
used a learning experiment in the fruit fly Drosophila which paralleled the operant concept proposed by Skinner: the tethered animals
first produce more or less random behaviors (including turning attempts, left
or right) and the experimenter rewards only designated correct ones until the
animal is spontaneously generating predominantly correct behaviors (e.g. left
turning attempts; Fig. 1). Importantly, we also used a control experiment, in
which the animals behavior not only affected whether they would receive the
reward or not, but also which color their environment was. Previous results had
shown that in this control situation flies tend to learn more about the coloration
of their environment than about their own behavior (3). If the function of dFoxP in flies were analogous to that of
FOXP2 in humans, we would expect it to be necessary for the first experiment
(operant self-learning), but not for the second experiment (operant
world-learning).
Ever since Skinners proposal,
these kinds of experiment had been discussed, but until now they have not been
technically feasible. In his critique of Skinners proposition, the linguist
Noam Chomsky dismissed the idea of operant experiments conceptually paralleling
language acquisition as mere homonyms, with at most a vague similarity of
meaning (4).
In order to be able to attribute
any effect of our manipulations in the flies to the dFoxP gene, we used two different strategies. In the first, we
tested flies with a mutation in the dFoxP gene in operant self- and world-learning. In the second we used the same two
experiments to test flies in which we had experimentally targeted the dFoxP gene such that its expression was
reduced. Both methods yielded essentially the same result: dFoxP is necessary for operant self-learning but not for operant
world-learning, lending support to the hypothesis that operant self-learning
may be one of the evolutionary ancestral capacities which had to exist in order
for language to be able to evolve (i.e., an exaptation).
dFoxP and habit formation
Another parallel between operant
and language learning is the fact that prolonged practice leads to an
automatization of the movements required. Only when a language is new do we
need to think about the pronunciation and articulation of words and sentences.
Once we are fluent, we only need to articulate our thoughts. Similarly, other
movements can be trained with feedback until they become automated. Riding a
bike, writing, tying shoe-laces, etc. are all examples of such automatic
behaviors called skills or habits. If the learning mechanism for which dFoxP is required constitutes an
exaptation for language acquisition and the speech component of language is a
special form of a skill or a habit, then dFoxP mutant flies should be deficient in habit formation. To test this hypothesis,
we used dFoxP mutant flies in a
prolonged operant world-learning paradigm known to induce habits (5). Further corroborating our
hypothesis, these mutant flies showed a severe deficit in habit formation.
Altered brain morphology in mutant flies
In vertebrate animals, mutations in
the FoxP2 gene leads to alterations in the brain structure of the affected
individuals (1). This is thought to be due to
the ability of FoxP genes to alter the expression of other genes, directly
involved in brain development. To test if the fly dFoxP gene also is involved in brain development, we reconstructed
the three-dimensional structure of the brains of flies with a mutated dFoxP gene in the computer. Using
computer-assisted volume analysis, we discovered alterations in the fly brain
structure which were too subtle to spot with the human eye, even at large
magnifications. These results indicate that in flies as in vertebrate animals,
FoxP genes may act as gene regulators during brain development.
Taken together our results provide
evidence for a structural and functional conservation of FoxP genes since the
split between vertebrate and invertebrate animals more than 500 million years ago. This deep homology spans
vastly different brain organizations.
Title of publication:
Drosophila FoxP mutants are deficient in operant self-learning
Source:
PLoS ONE: 10.1371/journal.pone.0100648
Authors:
EZEQUIEL MENDOZAa, JULIEN COLOMBb, JÜRGEN RYBAKc, HANS-JOACHIM PFLÜGERb, TROY ZARSd, CONSTANCE SCHARFFa, BJÖRN BREMBSb,e
Affiliations
a Inst. Biol. - Behavioral Biology, Freie Universität Berlin, Germany
b Inst. Biol. - Neurobiology, Freie Universität Berlin, Germany
c Neuroethology, Max-Planck Institute for Chemical Ecology, Jena, Germany
d Biological Sciences, University of Missouri, Columbia, USA
e Institut für Zoologie - Neurogenetik, Universität Regensburg, Germany
Corresponding author:
Björn Brembs
Universität Regensburg
Institute of Zoology - Neurogenetics
Universitätstr. 31
93053 Regensburg bjoern@brembs.net bjoern@brembs.net
+49-(0)941-943-3117
Abstract:
Intact function of the Forkhead Box P2 (FOXP2) gene is necessary for normal development of speech and language. This important role has recently been extended, first to other forms of vocal learning in animals and then also to other forms of motor learning. The homology in structure and in function among the FoxP gene members raises the possibility that the ancestral FoxP gene may have evolved as a crucial component of the neural circuitry mediating motor learning. Here we report that genetic manipulations of the single Drosophila orthologue, dFoxP, disrupt operant self-learning, a form of motor learning sharing several conceptually analogous features with language acquisition. Structural alterations of the dFoxP locus uncovered the role of dFoxP in operant self-learning and habit formation, as well as the dispensability of dFoxP for operant world-learning, in which no motor learning occurs. These manipulations also led to subtle alterations in the brain anatomy, including a reduced volume of the optic glomeruli. RNAi-mediated interference with dFoxP expression levels copied the behavioral phenotype of the mutant flies, even in the absence of mRNA degradation. Our results provide evidence that motor learning and language acquisition share a common ancestral trait still present in extant invertebrates, manifest in operant self-learning. This 'deep' homology probably traces back to before the split between vertebrate and invertebrate animals.
REFERENCES.
1. Bolhuis
JJ, Okanoya K, Scharff C (2010) Twitter evolution: converging mechanisms in
birdsong and human speech. Nature Reviews Neuroscience 11:747-759.
2. Skinner BF (1957) Verbal
Behavior (Copley Publishing Group).
3. Brembs B, Plendl W (2008)
Double dissociation of pkc and ac manipulations on operant and classical
learning in drosophila. Current Biology 18:1168-1171.
4. Chomsky N (1959) A Review
of B. F. Skinners Verbal Behavior. Language 35:26-58. Available at:
http://cogprints.org/1148.
5. Brembs B (2009) Mushroom
bodies regulate habit formation in Drosophila. Current Biology 19:1351-1355
|




